La Validamicina A (Val-A) es un agro-antibiótico
antimicótico (o antifúngico), producido por la bacteria Streptomyces hygroscopicus 5008 en la fermentación industrial y muy
utilizado, especialmente en Asia, para la protección de plantas contra
enfermedades producidas por Rhizoctonia.
Para la determinación de su producción se utiliza el
análisis de cromatografía líquida de alta resolución (HPLC). Esta técnica
permite separar los componentes de una mezcla basándose en las diferentes
interacciones químicas entre la sustancia analizada y la columna
cromatográfica. Sin embargo, este y los otros métodos que hoy se conocen,
presentan inconvenientes cuando se quieren analizar muestras grandes.
Actualmente, se está intentando desarrollar un nuevo
método basado en espectrofotometría (medición de la cantidad de energía
radiante que absorbe un sistema químico en función de la longitud de onda) para
la determinación de Val-A ya que, cuando esta es sintetizada, es secretado
simultáneamente el pigmento melanina. Además, se observa una relación estable entre la
concentración de Val-A y el valor de absorción espectral (SA) del pigmento a
450 nm. Por tanto, el método espectrofotométrico permite determinar la
producción de Val-A de forma rápida y sencilla.
Para comprobar su validez, se comparó la productividad
de Val-A por el método de HPLC con el método de SA y, con este último, se observaron resultados positivos que
permitían encontrar con éxito cepas con alta productividad de Val-A y tiempo
corto de fermentación.
Además, en muchas cepas el antibiótico y el pigmento
son regulados por el mismo factor y sus producciones están relacionadas. Por
tanto, en la fermentación de Streptomyces
hygroscopicus 5008 se podría establecer una ecuación basada en la relación
entre el valor de la absorción espectral del pigmento y la concentración de
Val-A en el caldo.
Por tanto, en este trabajo lo que se pretende es establecer
un método preciso, rápido y barato utilizando un medio industrial y una cepa de
fermentación para cuantificar la producción de ValA y, el método
espectrofotométrico anteriormente descrito, podría ser una buena alternativa.
Igualmente, esta investigación también
puede ser aprovechada para encontrar antibióticos en aquellas cepas productoras
de pigmento.
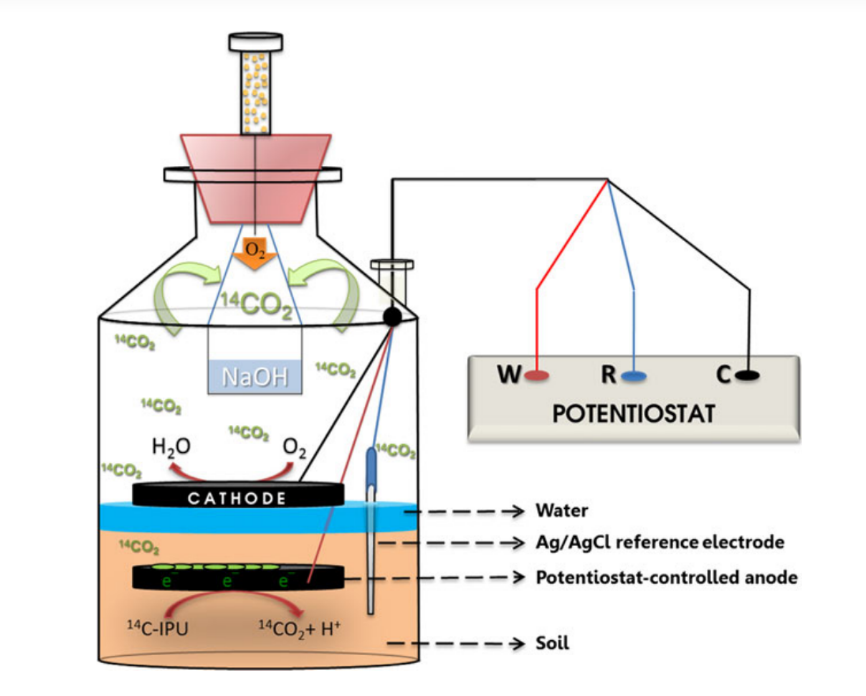
Para llevar a cabo el experimento se cultivó la cepa S. Hygroscopicus 5008 en placas de agar
con harina de soja y maltosa. Se recogieron las esporas desarrolladas y se
suspendieron en glicerol durante 7 días a 37ºC. Dicha suspensión de esporas fue
cultivada y posteriormente fermentada.
Por otro lado, había que comprobar si el pigmento
producido durante la biosíntesis de Val-A era estable. Se examinó su
estabilidad térmica, de la luz y la capacidad de resistencia al medio alcalino.
A continuación, se comparó el valor de absorción espectral de las muestras de
pigmento almacenado en las distintas condiciones.
El siguiente paso fue la medición de la producción de
Val-A. Para ello se tomaba una alícuota del caldo de fermentación cada 24 horas
a las que se sometía a posterior centrifugación. Se mezclaba el sobrenadante
resultante con cloroformo y, la fase acuosa superior la cual contenía
Val-A y el pigmento, se filtraba desechando la fase
orgánica inferior que presentaba proteínas y otros contaminantes. Este
sobrenadante filtrado se pasa a un nuevo tubo para las siguientes pruebas.
A continuación, para llevar a cabo la validación del
método espectrofotométrico para la determinación de Val-A, se tomaron tres
muestras del tubo anteriormente mencionado cada 24 horas. El valor de SA del
pigmento secretado y la producción de Val-A detectados mediante HPCL se
ajustaron a una ecuación mediante el software STATA. Se probó que el
experimento era repetible y se correspondía con dicha ecuación.
Por otro lado, para eliminar el efecto de los
diferentes componentes del medio que puedan afectar en el color del caldo de
fermentación se seleccionaron tres medios: el medio A (descrito anteriormente),
el medio B (polvo de arroz, harina de soja, NaCl KH2PO4 y
CaCO3) y el medio C (almidón soluble, extracto de levadura, KH2PO4
y NaCl).
También se evaluó la aplicabilidad de la
espectrofotometría en la estimación de la producción de Val-A utilizando
muestras en condiciones de fermentación diferentes. Lo que se pretendía era
observar la relación entre el pigmento y la productividad de Val-A cuando se
añadía un estimulante exógeno, el cual se había demostrado que mejoraba la productividad
de Val-A. Dado que el pH es un factor importante que influye en el metabolismo
microbiano y en el color de pigmento se realizaron varios experimentos variando
el pH.
Por último, se generaron cepas mutantes (mediante
irradiación ultravioleta y N-metil-NO-nitroN-nitrosoguanidina) para seleccionar
aquellas cepas de Val-A que tuvieran alto rendimiento.
Se ha
estudiado la estabilidad del pigmento en diferentes condiciones anteriormente
descritas, por lo que el pigmento se relaciona con la producción de Val-A.
Como se muestra en la Fig. 1, el crecimiento de S. hygroscopicus 5008, junto con la producción de Val-A, condujo a una serie de cambios
regulares en el color del caldo de la fermentación. La producción de pigmento y
Val-A en condiciones óptimas de cultivo mejoraban ambos su producción, mientras
que en condiciones desfavorables, sus metabolismos eran suprimidos. En el
análisis transcripcional de los genes de biosíntesis de Val-A y la respuesta
del gen relacionado con el pigmento, se demostró que presentaban cooperatividad
en diferentes condiciones de fermentación, por tanto, ambos están regulados por
los mismos reguladores.

Fig 1. Cambio de
color del caldo de fermentación en la producción de Val-A.
Es posible determinar que existe una relación estable con la
producción de Val-A y la correlación entre el color del caldo, por lo que la
producción de Val-A podría ser la base del método de espectofotometría. Se escogió
el valor de 450 nm para medir la absorbancia de la melanina (pigmento), que
presentaba una relación constante a dicho valor, el cambio se producía debido a
la concentración de Val-A de las muestras. Esto llevó a que 450 nm se
estableciera como la longitud de onda de detección para establecer la fórmula
de ajuste de la producción de Val-A.
El siguiente paso es la determinación de la fórmula
de ajuste en el ensayo de espectofotometría. Para obtener la ecuación tomamos
como variables los valores de la concentración de Val-A (Y) determinados por
HPLC y el valor de SA a 450 nm (X):

El coeficiente de determinación de la ecuación es 0,9928, por lo que muestra un alto grado de ajuste. En la FIG 2., se muestra la curva de ajuste correspondiente a 40 muestras en diferentes cultivos o condiciones de fermentación.
El coeficiente de determinación de la ecuación es 0,9928, por lo que muestra un alto grado de ajuste. En la FIG 2., se muestra la curva de ajuste correspondiente a 40 muestras en diferentes cultivos o condiciones de fermentación.

FIG 2. Curva de
ajuste de la relación entre la producción de Val-A y el valor de DO (densidad
óptica) del pigmento
Los resultados en unas condiciones de fermentación
deben ser verificados por test para una probabilidad dada, normalmente p=0,05.
Los ensayos de este trabajo han sido validados por el t-test. El método de HPLC
para la determinación de Val-A ha sido ya verificado, por lo que se comparó con
el método SA a este.

En la Tabla 1, a diferentes tiempos de fermentación
todos los valores de p son superiores a 0,05 (2ª columna), por lo que en el
intervalo de confianza del 95% no hay diferencia entre las concentraciones de
Val-A detectadas por HPLC y por SA.
El mayor R.S.D (Desviación Estándar Relativa) en SA es
de 0.212 correspondiente a las 48 h, al no ser en este momento el de mayor
secreción de Val-A ni de pigmento, esta desviación puede ser aceptada. A las 96
h, es el momento de máxima producción de Val-A. Las R.S.D correspondientes a
HPLC y SA a este tiempo son 0.027 y 0.007 respectivamente, lo que confirma
que el método SA tiene una buena estabilidad al igual que el método HPLC.
En los ensayos para la verificación de la viabilidad
del método SA se llegó a conclusiones tales como que la productividad de Val-A
es mayor a una Tª relativamente más alta junto a una síntesis de proteínas y
consumo de azúcar más rápidos. Para verificar la exactitud, repetibilidad y
aplicabilidad de la ecuación de ajuste para la determinación cuantitativa
rápida de Val-A, se compararon diferentes medios y condiciones de fermentación,
tales como la adición de gbutirolactona, etanol, pH, etc. Se comparó una vez
más con el método HPLC quedando recogido en la Tabla 2. Además, se analizó la
precisión de la espectofotometría a las 120 h, tiempo en el que finaliza la
fermentación. Todo mediante la t-test.

Si tanto el valor Pr (T <t) como el de Pr (T> t)
[Pr (T> t)=1-Pr (T <t)] son superiores a 0,05, se considera que la
hipótesis es aceptada. El resultado demostró que no existía ninguna desviación
del paralelismo en este estudio, por lo tanto, el método SA establecido podría
considerarse tan exacto como el método HPLC. Aunque los cambios en los cultivos
y condiciones de fermentación afectaran a la producción de Val-A y el
metabolismo de S. hygroscopicus 5008,
se mantenía la relación entre Val-A y el
valor SA, por lo que no afecta a la exactitud y estabilidad de los valores
predictivos calculados por la ecuación de ajuste.
El pretratamiento de muestras no introduce sustancias
que puedan afectar a la DO del caldo a 450 nm y el pigmento puede ser estable
en un entorno común durante mucho tiempo. Como el método SA se basa en la
densidad óptica del pigmento, podría utilizarse para investigar la relación
entre la producción de antibióticos y la secreción de pigmentos desde la
perspectiva del metabolismo microbiano. Siempre que se
pueda obtener una relación cuantitativa y estable, el método SA se podrá
utilizar para cuantificar otros antibióticos en la fermentación.
La cuantificación de Val-A por método de HPLC es
precisa, pero el equipo y los reactivos relacionados para HPLC cuestan mucho.
El tiempo de análisis para una muestra es de unos 20 minutos, sin incluir la
preparación de reactivos, requiriendo todo esto para un gran número de
muestras. Todo esto hace que no sea un buen método para la fermentación
industrializada, en contra, el método SA no necesita instrumental y reactivos
costosos, y además, una gran cantidad de muestras pueden ser analizadas en la
placa de 96 pocillos a la vez.

FIG 3. Cribado de mutantes de alto rendimiento por el método
SA en placa de 96 pocillos y análisis comparativo de la producción de Val-A
mediante método SA y HPLC. Cinco días cultivo wild type 5008 y siete mutantes
en dos medios diferentes. (A) La producción de Val-A en el quinto día por el
método SA y por HPLC representado en gráfico de barras (B).
El método de ensayo SA descrito en placa de 96
pocillos se utilizó para determinar la productividad de Val-A y el cribado de
alto rendimiento (Fig. 3). Siete
mutantes se compararon con los de tipo salvaje 5008 (WT). La productividad de
Val-A por el método de SA fue comparada con el método de HPLC. El mutante 4
(M4) fue determinado como la cepa positiva con la mayor productividad (Figura
3B). Además de una selección rápida y conveniente de Cepas, el tiempo de
fermentación de los mutantes puede estimarse utilizando SA como un método de
monitorización rápido en tiempo real Val-A. También es posible emplear el
método SA para cepas con tiempos de fermentación cortos, por lo que es un
método con amplias posibilidades de aplicación. Por ahora, no existe un método
de prueba rápido de Val-A que pueda satisfacer la demanda industrial ni el
ensayo SA en la determinación cuantitativa de antibióticos.
Referencia bibliográfica:
http://www.sciencedirect.com/science/article/pii/S1389172316301153?via%3Dihub
Más información acerca de Rhizoctonia:
http://www.cyclamen.com/es/profesional/enfermedades/8/22
Nombres:
Gracia Patricia Mínguez Marco
Carmen Mateu Olivares
Ana Mª Rodríguez Lozoya
Alba Mª Carrascosa Costa
Laura Serrano Arratia
Carmen Molina Pardines
Referencia bibliográfica:
http://www.sciencedirect.com/science/article/pii/S1389172316301153?via%3Dihub
Más información acerca de Rhizoctonia:
http://www.cyclamen.com/es/profesional/enfermedades/8/22
Nombres:
Gracia Patricia Mínguez Marco
Carmen Mateu Olivares
Ana Mª Rodríguez Lozoya
Alba Mª Carrascosa Costa
Laura Serrano Arratia
Carmen Molina Pardines

















{kind=link}
{kind=link}
{kind=link}